11. De la Variación a la Especiación, Parte 1
16 de Mayo de 2013. Temas: Genética
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo tratamos sobre cómo se extiende la variación en la población, y cómo pueden surgir, con el tiempo, las diferencias entre las poblaciones.
En el último artículo de esta serie examinábamos cómo la variación en el ADN surge por sucesos aleatorios tales como los fallos en el emparejamiento de las bases, las duplicaciones y las deleciones. Para entender cómo esta variación puede, finalmente, contribuir a la especiación, tenemos que discutir cómo la variación se propaga por la población. En primer lugar, necesitamos algo de vocabulario para facilitar la discusión: tenemos que explicar, específicamente, la diferencia entre un gen y un alelo.
Como genético, me tiro de los pelos, pierdo la paciencia, a veces, al leer en los medios informes popularizados sobre temas científicos. Una de las cosas que más me molesta es la utilización de la palabra “gen” para decir que un individuo “tiene un gen” para un rasgo específico. Ya hemos explicado que los genes son una parte de la secuencia de ADN de un cromosoma que contribuye a una función de algún tipo, generalmente codificando un producto proteínico. En este sentido, los hombres tienen todos los mismos genes, o casi: cerca de 20 000 secuencias, más o menos, que biológicamente nos hacen como somos. Lo que, sin embargo, no tenemos son genes idénticos: hay diferencias que surgen de los errores de copia que ya hemos discutido. Estas diferencias se llaman alelos. Se puede pensar en un alelo como en una “versión” o como un “sabor” alternativos de un gen. Las mutaciones no suelen crear genes nuevos, aunque pueden hacerlo a través de la duplicación. Generalmente, lo que se crean son alelos nuevos. En un artículo precedente, utilizábamos las piezas de las construcciones infantiles para ilustrar cómo puede surgir una variante nueva a través de un fallo en el emparejamiento de las bases del ADN durante la replicación cromosómica:
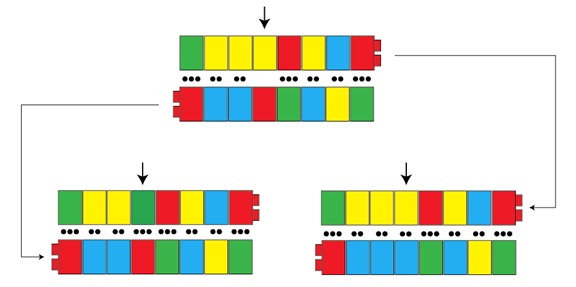
En este ejemplo, tenemos una secuencia que, a través de un error de copia, se convierte en dos versiones ligeramente diferentes de lo que es, casi, la misma secuencia. Estas diferencias son lo que se llamarían dos alelos distintos y, si afectaran a la función de un gen, podrían tener un efecto detectable a nivel del organismo en su conjunto. Cuando los medios hablan del “gen” para esta o aquella característica, a lo que realmente se refieren es a un alelo de una característica dada: a la variante específica de un gen que se correlaciona con una determinada condición médica, por ejemplo.
Selección y deriva
Así pues, la variación en el ADN es precisamente la producción de los alelos nuevos, pero ¿qué pasa con estos alelos, a lo largo del tiempo, dentro de una población? Obviamente, cuando surge un alelo nuevo, está presente sólo en un individuo. Para que tenga algún impacto sobre el conjunto de la población tiene que propagarse a otros individuos pasándolo a las crías. De esta forma, la variación puede entrar en una población y, con el tiempo, hacerse más común. Hay una serie de factores que pueden influir en este proceso. Si el tamaño de la población es pequeño, el propio azar puede hacer que aumente, o disminuya, la frecuencia de un alelo en la población, lo que se conoce como deriva genética. Como la deriva puede ser un actor principal, en cuanto al modo en que las frecuencias alélicas cambian con el tiempo en una población, vale la pena emplear algún tiempo para discutirlo con cierto detalle.
La deriva es esencial en un muestreo no representativo. Consideremos una pequeña población de organismos con reproducción sexual, que podemos representar con rectángulos, cada uno de los cuales contiene dos alelos (los cuadrados; siendo los colores las diferencias alélicas). Podemos representar su “reproducción” como con una probabilidad del 50% (50:50) de pasar cada uno de los dos alelos a las crías de la generación siguiente. (Nota: cada suceso de “paso a las crías” es independiente del resto. Por ejemplo: No hay mecanismo que garantice que un individuo pueda pasar sus dos alelos, reproduciéndose dos veces.) En la pareja reproductora de la izquierda, uno de los padres tiene dos alelos amarillos, y el otro tiene un alelo azul y otro alelo amarillo. Cuando se reproducen, el padre con los dos alelos amarillos pasa, por azar, sólo el alelo amarillo a sus dos crías. En la pareja de reproductores de la izquierda, el padre con los dos alelos, amarillo y azul pasa, por azar, el alelo amarillo dos veces, y el azul ninguna.
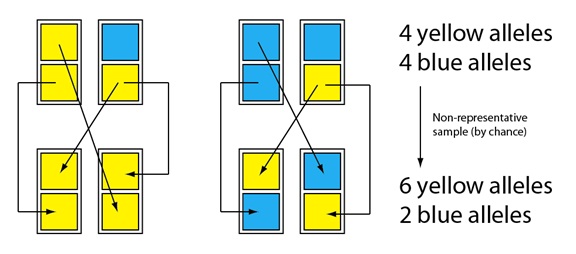
4 alelos amarillos
4 alelos azules
Muestra no representativa
(por azar)
6 alelos amarillos
2 alelos azules
Imaginemos ahora que las crías se emparejan para reproducirse y que, de nuevo tenemos, por azar, una muestra ligeramente no representativa para formar la siguiente generación:
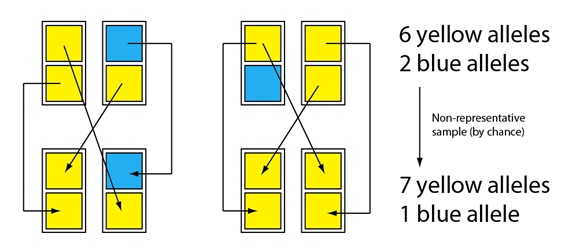
6 alelos amarillos
2 alelos azules
Muestra no representativa
(por azar)
7 alelos amarillos
1 alelo azul
El problema es que esta pequeña población es propensa a sufrir amplias fluctuaciones en las frecuencias de los alelos azul o amarillo por lo pequeña que es. Este pequeño tamaño significa que los sucesos aleatorios, incluso con una sola pareja reproductora, tiene un gran impacto en la población en su conjunto. En la generación siguiente, por ejemplo, el alelo azul podría perderse por completo; y una vez que desapareciera permanecería ausente hasta que, o bien reapareciera por una nueva mutación, o bien entre en la población a través de la migración de un individuo que venga de una población en la que sí está presente.
En poblaciones grandes, sin embargo, la situación es bastante diferente. Imaginemos una población de 1000 individuos, con un total de 500 alelos azules y 500 amarillos repartidos aleatoriamente entre los individuos. Cuando esta población se reproduce, nunca variará demasiado esa proporción del 50% (50:50) de una generación a otra. En esta población grande, los sucesos aleatorios en una pareja de reproductores son, proporcionalmente, sólo una pequeña parte de la población en su conjunto y, como valor medio, la población seguirá reflejando la probabilidad de un 50% (50:50) de que cualquier alelo pase a la siguiente generación.
Así que, ¿Cómo pueden cambiar las frecuencias alélicas en poblaciones grandes, cuando la deriva no puede hacerlo? Ya hemos visto un mecanismo que puede lograrlo: la selección natural. La selección natural es simplemente el efecto de que los individuos que tienen un alelo se reproduzcan con mayor frecuencia que los que no tienen ese alelo. Con el tiempo, este sesgo de la probabilidad de reproducción aumenta la frecuencia de los alelos seleccionados en la población. En el perro, en los albores del proceso de domesticación, la duplicación de los genes de la amilasa sucedió como una mutación aleatoria, una única vez. Los perros que tenían el alelo duplicado de la amilasa se reprodujeron a un ritmo ligeramente mayor que los que no, porque el alelo duplicado les permitía nutrirse mejor con el alimento que recibían en su nuevo ambiente: los recursos humanos. Con el tiempo, el alelo duplicado se volvió tan frecuente en la población de perros que el alelo ancestral, no duplicado, se perdió del todo. En ese punto el alelo nuevo se “fijó” en la población: alcanzó una frecuencia del 100%.
Resumiendo: en poblaciones pequeñas, la deriva puede tener un gran impacto sobre las frecuencias alélicas entre una generación y la siguiente. En poblaciones grandes, predomina la selección natural, y la deriva tiene un impacto reducido. Ambos mecanismos pueden contribuir al cambio de las frecuencias alélicas en la población a lo largo del tiempo y, como tales, ambos pueden ser factores que contribuyen a la especiación.
El cambio de frecuencias alélicas y la especiación
Especiación es la producción de especies nuevas a partir de una población ancestral. (Pero ya hemos discutido cómo la definición de “especie” resulta un concepto confuso, poco claro, y es el hecho de que surjan lenta y progresivamente lo que les hace difíciles de definir).
Una forma de entender cómo empieza la especiación es considerar dos poblaciones de la misma especie que, por alguna razón, dejen de reproducirse entre ellas; quizá a través del aislamiento geográfico. Aunque una “barrera geográfica” no tiene nada que ver con las diferencias genéticas o la compatibilidad reproductiva, si tal barrera existe, los alelos que surgen en una población no serán transferidos a la otra. Además, si dos poblaciones no están intercambiando alelos, las frecuencias alélicas en las dos poblaciones ya no estarán ligadas entre sí ni promediadas entre ellas. Ello significa que la deriva y la selección van a actuar ahora independientemente en las dos poblaciones. Una vez desacopladas, las dos poblaciones pueden seguir trayectorias diferentes; una población puede comenzar siendo pequeña, y ser dominada por la deriva hasta que aumenta su tamaño. La otra población puede seguir siendo grande, y estar sometida a la selección natural de formas en que la otra población no lo es. En un período de tiempo amplio, las dos poblaciones pueden llegar a ser lo bastante diferentes, genéticamente, como para constituir dos especies distintas. La clave, desde luego, es la naturaleza de la barrera que impide el intercambio de alelos entre las poblaciones. En el siguiente artículo de esta serie examinaremos cómo pueden aparecer esas barreras en las poblaciones.


