12. De la Variación a la Especiación, Parte 2
17 de Mayo de 2013. Temas: Genética
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo tratamos sobre cómo surgen las barreras geográficas que limitan el flujo de alelos entre las poblaciones, y cómo el efecto fundador puede contribuir a las diferencias genéticas en las poblaciones recién fundadas.
Repaso
En el artículo de ayer establecíamos una serie de puntos que vale la pena resumir aquí:
Los alelos nuevos surgen como sucesos únicos en los individuos, pero pueden volverse comunes en la población a través de varios procesos, incluidos la deriva genética y la selección natural.
Los nuevos alelos, de volverse comunes en la población, pueden variar los caracteres promedio de la misma.
Si, entre dos poblaciones de una misma especie, el intercambio de alelos queda bloqueado o reducido, las características medias de las dos poblaciones pueden divergir entre sí.
Con el tiempo suficiente, estos procesos pueden dar lugar a diferencias entre los dos grupos lo suficientemente significativas como para convertirlas en especies diferentes.
Con esta serie de puntos en mente, podemos ahora ver con más detalle las diversas formas en que el intercambio genético entre poblaciones puede ser reducido o incluso eliminado. Empezaremos con el caso más simple, el del aislamiento geográfico completo.
Barreras geográficas
La separación geográfica de dos poblaciones de una misma especie es una manera rápida y efectiva de detener el intercambio de alelos entre ellas. En el momento de la separación las dos poblaciones son, desde luego, del todo capaces de cruzarse entre sí, pero la separación física les impide hacerlo. Un ejemplo de aislamiento geográfico que conduce a la especiación, y que ya hemos discutido con anterioridad, es el de las diversas especies de pinzones que Darwin observó en las islas Galápagos de la costa de Suramérica. La población original de pinzones de las Galápagos fue fundada por un pequeño grupo de aves que llegaron a las islas procedentes del área continental de Suramérica, muy probablemente arrastradas allí durante una tormenta. Estas aves, como población, quedaron biológicamente aisladas de su población de origen en el continente, ya que las Galápagos están a cientos de kilómetros mar adentro. Una vez separado de la población mayor, el pequeño grupo “fundador” dejó de recibir nuevos alelos de ella, y tampoco volvió a pasar de vuelta los nuevos alelos que surgieran en él. A pesar de tratarse de dos poblaciones de la misma especie, ahora estaban genéticamente aisladas una de la otra, y las diferencias en las frecuencias alélicas entre ellas comenzaron a acumularse. Estas diferencias, con el tiempo, dieron lugar a cambios en las características medias y, finalmente, a la formación de una especie nueva.
El efecto fundador
En muchos casos este proceso de acumulación de diferencias tiene su comienzo en el mismo momento de la separación, por el fenómeno conocido como “efecto fundador”. Una pequeña población fundadora es, muy a menudo, una muestra no representativa de la diversidad genética de la población de origen. Por ejemplo, consideremos una población hipotética de 36 individuos. Cada individuo lleva dos alelos de un determinado gen, y en la población hay cuatro alelos diferentes de ese gen, representados aquí por los cuatro colores:
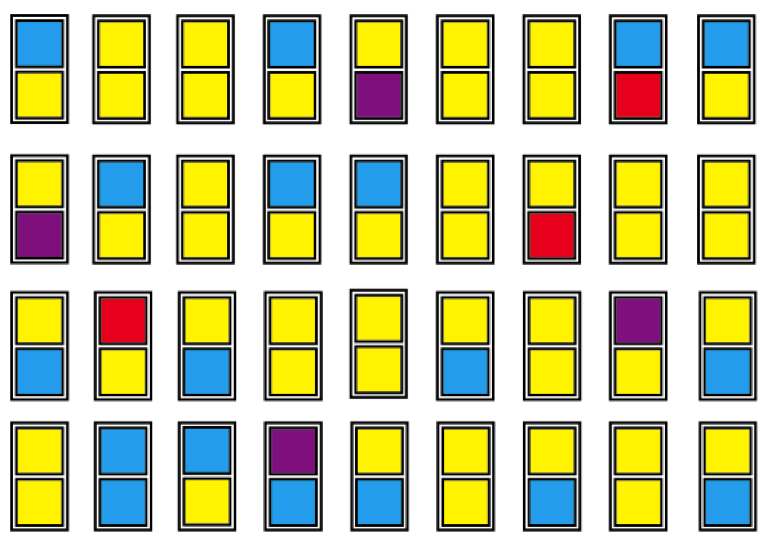
Observemos que el alelo amarillo es el más común, seguido del azul. Los alelos púrpura y rojo son relativamente raros en esta población. De hecho, su rareza indica que será muy improbable que apareciera un individuo con dos alelos rojos en la generación siguiente, por ejemplo. Para lograr tal individuo, habrían tenido que emparejarse dos padres que fueran “portadores” del alelo rojo, y habrían tenido que pasar ambos ese alelo rojo a la cría. Esto, aunque no es imposible, en esta población sería improbable.
Ahora supongamos que algunos miembros de esta población empiezan una nueva en una isla aislada. Sólo seis individuos forman la nueva población, y los alelos que ellos llevan no constituyen una representación perfecta de las frecuencias alélicas de la población de origen, mucho mayor (se muestran las frecuencias aproximadas en la población de origen y en la nueva población “fundadora”):
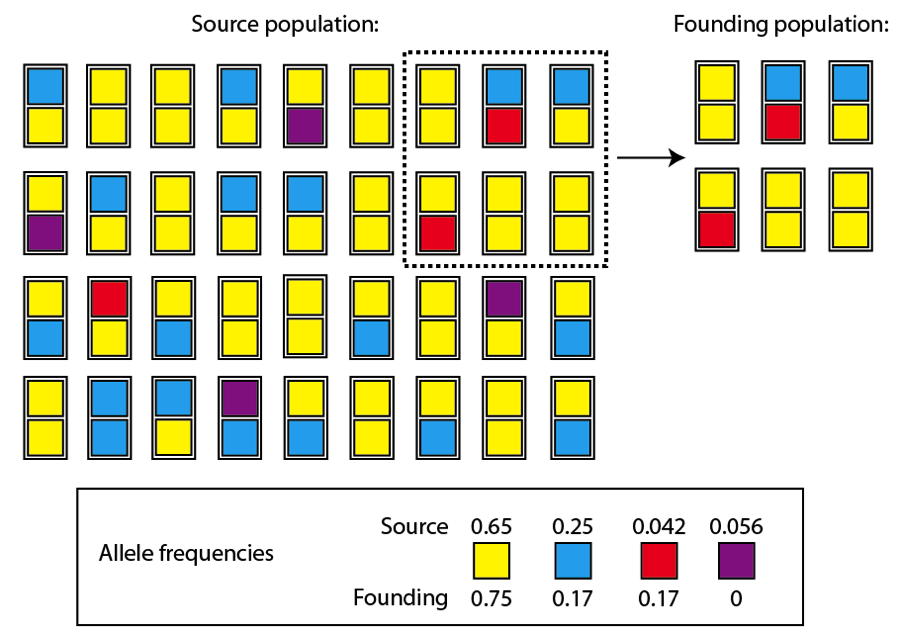
Población de origen Población “fundadora”
Origen 0.65…
Frecuencias alélicas
Fundadora 0.75…
Podemos ver que, entre los alelos comunes, la frecuencia del amarillo ha aumentado y la del azul ha disminuido. A pesar de estas diferencias, la frecuencia de los alelos comunes es razonablemente similar a la de la población de origen. Los alelos raros, sin embargo, han sufrido mayores cambios: el alelo rojo es ahora mucho más común en la población recién fundada y el púrpura ha desaparecido por completo.
Pues bien, estos cambios son sutiles y los cambios en un gen no es probable que sean suficientes como para precipitar la especiación entre las dos poblaciones. Sin embargo, estos tipos de cambios podrían ser significativos a largo plazo. Pensemos en el alelo rojo en la población recién fundada. A medida que esta población aumente en número, será mucho más probable que aparezcan individuos con sus dos alelos rojos en esta población que en la población de origen. Si esta combinación genética tiene una ventaja selectiva, la selección natural podrá actuar sobre ella en la nueva población. Sin embargo, en la población de origen esta combinación genética es mucho más improbable, volviendo muy difícil la actuación de la selección natural sobre esta combinación de alelos. Con el tiempo, el alelo rojo podría llegar a ser dominante en la población nueva, pero seguirá siendo raro en la de origen. Además, es probable que el ambiente sea algo distinto para estas dos poblaciones, lo que producirá diferencias en la selección natural. Lo que pudiera ser una combinación alélica ganadora en el área continental, podría no ser tan adecuada en el ambiente insular, y viceversa. Otro tema es que la población recién fundada, como cualquier población pequeña, está mucho más sometida a la deriva genética que la población de origen, que es mayor. El alelo rojo podría aumentar su frecuencia en la nueva población simplemente por azar y no por la acción de la selección natural.
En conjunto, estos mecanismos pueden situar a las dos poblaciones en trayectorias distintas y, con el tiempo, producir diferencias significativas entre ellas. Con tiempo suficiente, las diferencias acumuladas pueden ser suficientes como para mantener las poblaciones separadas, incluso si éstas volvieran a entrar en contacto. En tal caso, los biólogos clasificarían a las dos poblaciones como especies distintas. Aunque esto resulta más fácil con especies que han permanecido separadas mucho tiempo, que han acumulado diferencias significativas y que, por lo tanto, ya no se cruzan entre sí, o lo hacen muy esporádicamente, resulta realmente difícil con poblaciones separadas recientemente y que no están todavía reproductivamente aisladas. Por lo tanto, lo que constituye una “verdadera especie” y no simplemente una “subespecie” o una “variedad” es, a menudo, objeto de discusión y debate entre científicos y, de hecho, fue un tema al que Darwin dedicó mucho tiempo en sus trabajos. La ambigüedad surge del mecanismo de la divergencia, lenta y gradual, de las especies a partir de una población ancestral común.
No sólo diferencias
Dado todo lo dicho anteriormente, podríamos tener la impresión de que las diferencias entre las especies son el tema principal. Es cierto que las diferencias son vitales porque, a fin de cuentas, será la acumulación de diferencias lo que llevará a la formación de una especie nueva. Sin embargo, es importante recordar que, en especies estrechamente relacionadas, estas diferencias serán pocas si se comparan con las características que permanecen inalteradas en ambos grupos. A nivel genético, podemos ilustrarlo considerando un gen para el que sólo existiera un alelo en la población de origen; quizá un alelo que ya haya estado sometido a la selección natural y haya desplazado a todos los otros alelos. La población recién fundada heredará sólo este alelo, a pesar de la pequeña muestra del grupo fundador, porque no hay otras variantes en la población. El resultado será que la población insular es, para este rasgo, idéntica a la del continente hasta que una mutación, en cualquiera de las dos poblaciones, pueda plantear la posibilidad de un cambio. En la mayoría de los rasgos no surgirán mutaciones, porque los mecanismos de copia del ADN son muy precisos. Esto mantendrá constantes la mayoría de los rasgos entre las dos poblaciones. El patrón que esperamos para especies recientemente divergentes es, por tanto, de características mayoritariamente idénticas, sobre las cuales destacan unas pocas diferencias. Recordaremos que fue precisamente este modelo el que, en su contexto biogeográfico, le hizo a Darwin reflexionar sobre la posibilidad de que las especies pudieran no ser estables:
“Para nosotros el hecho más importante y llamativo, en relación con los habitantes de las islas, es su afinidad con los del continente más cercano, sin ser en realidad las mismas especies. Se podrían dar numerosos ejemplos de este hecho. Voy a dar solamente uno, el del archipiélago de las Galápagos, situado en el Ecuador, a 500 ó 600 millas de las costas de Sudamérica. Aquí, casi cada especie terrestre o acuática lleva el sello inconfundible del continente suramericano. Hay veintiséis formas de aves terrestres, veinticinco de las cuales son clasificadas como especies diferentes por el Sr. Gould, supuestamente creadas aquí; pero la gran afinidad de la mayoría de estas aves con especies americanas, en cada carácter, en sus hábitos, gestos y tonos de voz, resultaba manifiesta.
Destaquemos que fue la combinación de las “abrumadoras” similitudes, es decir características distintivas en común, con las sutiles pero significativas diferencias, lo que Darwin observó. Las aves en cuestión eran especies distintas, pero retenían el “sello inconfundible” de su herencia. Estas observaciones son las que llevaron a Darwin a plantearse la hipótesis de que estas especies de pinzones fueran el producto de una especiación producida a través del aislamiento geográfico.
Mientras que el aislamiento geográfico es una situación que directamente puede constituir barreras genéticas y dar lugar a la formación de especies nuevas, la especiación puede también suceder sin una separación completa. En el siguiente artículo de esta serie vamos a examinar un caso de especiación, con una barrera geográfica, y genética, sólo parcial; un caso que demostrará también lo “confuso”, lo poco claro, que resulta precisar exactamente lo que constituye una especie.


