13. De la Variación a la Especiación, Parte 3
30 de Mayo de 2013. Temas: Genética, Historia de la Vida
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo examinamos la difusa línea que existe entre especies y subespecies utilizando las llamadas “especies en anillo”, y discutiremos la especiación basada en el reparto de los recursos.
En el último artículo de esta serie veíamos el caso relativamente sencillo de la separación geográfica entre poblaciones, como puede ser la fundación de una nueva población en una isla. La separación geográfica es una barrera efectiva para lo que los biólogos llaman el “flujo de genes” entre poblaciones; un efecto más propiamente descrito como “flujo de alelos”. A medida que surgen nuevos alelos en poblaciones separadas, la falta de cruzamiento entre ellas mantiene a cada alelo en la población en que surgió. Los nuevos alelos pueden, con el tiempo, contribuir a la especiación si afectan a las características del organismo. Si, por el contrario, los alelos nuevos pueden pasar libremente entre las dos poblaciones entonces no contribuirán a dicha especiación, porque no estarán haciendo que las dos poblaciones se diferencien más con el tiempo.
Lo que se va, acaba volviendo1
Aunque estos dos extremos (poblaciones geográficamente separadas y poblaciones del todo continuas) se entienden fácilmente, es posible encontrar situaciones intermedias que son como los grises, entre el blanco y el negro. Por ejemplo, imaginemos dos poblaciones, que llamaremos “A” y “B”, y que pertenecen a una misma especie. Pueden intercambiar alelos entre ellas pero a un ritmo reducido en comparación con el ritmo con el que lo hacen dentro de cada una de ellas. Este efecto puede originarse a causa de la forma geográfica de su hábitat: Si éste es largo y estrecho, las dos poblaciones pueden colindar, una con la otra, sólo en una pequeña parte de su rango. Esto significa que, por término medio, un individuo de la población A es más probable que encuentre pareja dentro de la población A que se aparee con un miembro de la población B en su pequeña área de solapamiento. Podemos ilustrar esto con cajas que representen a las dos poblaciones colindando entre sí a lo largo de uno de sus lados cortos:
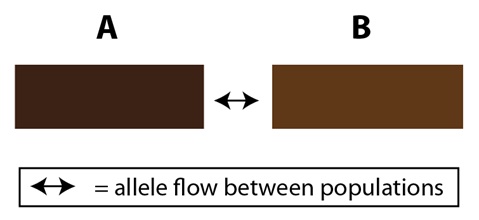
<—> = flujo de alelos entre las poblaciones
Esta disposición restringe por tanto, pero no impide completamente, el flujo de alelos entre las dos poblaciones. En efecto, esto es una barrera parcial al flujo de alelos. Las poblaciones A y B son miembros de la misma especie, pero no son las dos genéticamente idénticas. Cuando surgen nuevos alelos en la población A, ya no son compartidos con la población B con tanta frecuencia como lo son dentro de la A, y viceversa. Como tales, las poblaciones A y B pueden tener distintas frecuencias de un determinado alelo y una puede incluso tener algunos alelos que la otra no tiene. Igualmente es posible que las dos poblaciones experimenten diferencias en la selección natural ya que sus ambientes no son idénticos; y también puede haber diferencias en la deriva genética dependiendo del tamaño de cada una de las poblaciones. El resultado final es un equilibrio de las fuerzas que actúan sobre las dos poblaciones; unas favoreciendo las diferencias (selección y/o deriva) y otras favoreciendo las similitudes (flujo de alelos limitado a través de los cruzamientos).
En la naturaleza, este efecto puede extenderse a múltiples poblaciones en una cadena que se prolonga a través de una franja de hábitat apropiado. Añadamos al ejemplo anterior tres poblaciones más, C, D y E, para ilustrarlo:
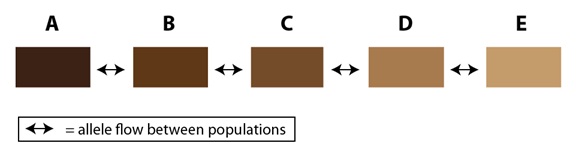
<—> = flujo de alelos entre las poblaciones
Una vez que las poblaciones se extienden por una amplia área geográfica, las diferencias entre las poblaciones en los extremos (poblaciones A y E, en el diagrama) pueden llegar a ser bastante significativas. En algunos casos, curiosamente, las poblaciones en los extremos de esa cadena pueden ser lo bastante diferentes como para no reconocerse entre sí como miembros de la misma especie, a pesar del hecho de estar genéticamente conectadas a través de una serie de poblaciones intermedias. En algunos casos, los científicos tienen que llegar a poner a miembros de las poblaciones de los extremos en contacto para ver si son capaces de cruzarse entre sí (es decir, utilizando el concepto biológico de especie como definición de especie). En otros casos, la topografía del hábitat los vuelve a reunir en la naturaleza, haciendo posible el reencuentro entre las poblaciones de los extremos de la cadena alrededor de un anillo, pero con una barrera natural en medio (como puedan ser una montaña o un valle con hábitat inadecuado). El resultado es lo que conocemos como “especies en anillo”:
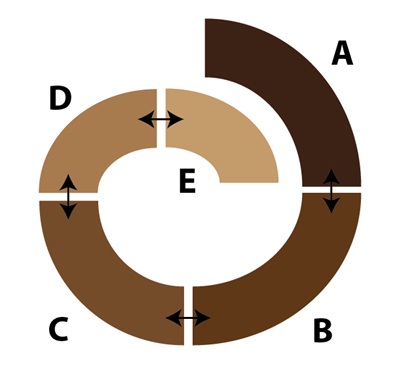
Se puede ver así lo difícil que resulta el definir qué poblaciones son especies separadas distintas (si es que algunas lo son). Hay flujo de alelos entre todas las poblaciones, pero sólo alrededor del anillo. Las dos poblaciones de los extremos, en la zona de solapamiento, a pesar de reencontrarse en el mismo hábitat, son lo bastante diferentes como para no cruzarse entre sí. Definir, o no, estas poblaciones como especies separadas es un intento infructuoso de trazar una línea de demarcación en lo que en realidad es un gradiente. Para los que estén interesados en un ejemplo real de una especie en anillo, las subespecies de la salamandra Ensatina eschscholtzii en la costa oeste Norteamericana son tanto un ejemplo de libro de texto como un tema de investigación en marcha.
Ahora bien, si encontráramos las poblaciones de los extremos en la naturaleza sin las formas intermedias, sin las poblaciones “puente”, no dudaríamos en clasificarlas como especies distintas. También es fácil ver lo que sucedería si se perdiera cualquiera de las poblaciones puente, o si los cambios en el hábitat interrumpieran la conexión con cualquiera de ellas. El resultado sería una rotura en la cadena de flujo de los alelos, desconectando a las poblaciones terminales una de la otra. Lo que ilustran las especies en anillo es que la especiación, aunque es un proceso lento de acumulación de diferencias entre poblaciones, también es posible incluso sin que haya una barrera completa al flujo de alelos.
Especiación sin separación geográfica
Aunque las especies en anillo ilustran cómo las especies pueden formarse produciéndose fragmentaciones en la variación a lo largo de una amplia área geográfica, también es posible que surjan barreras al flujo de alelos dentro de la población en una región geográficamente más compacta. Todo lo que hace falta es un sesgo que promueva el intercambio de alelos dentro de un subgrupo de la población a expensas del intercambio con la población general más amplia; y, como hemos visto en el caso de las especies en anillo, esta barrera no necesita ser absoluta para permitir que dos subpoblaciones acumulen diferencias y diverjan una de la otra con el tiempo. Una forma de lograrlo es que las subpoblaciones empiecen a explotar los recursos de un área geográfica común de distinta manera unas de otras; un efecto conocido como reparto de recursos. Cuando las subpoblaciones empiezan a especializarse en “formas de vida” ligeramente distintas, como dijo Darwin, se vuelven más proclives a cruzarse dentro de su subpoblación que con la población en su conjunto. Como la reproducción preferencial es una barrera (parcial) al flujo de alelos, esto puede colocar a las dos subpoblaciones en una trayectoria genética que refuerce sus diferencias y conduzca a una especiación. El reparto de recursos es el probable mecanismo que dirige los múltiples y rápidos sucesos de especiación que tienen lugar cuando una población fundadora llega a un nuevo hábitat en el que los competidores están, en general, ausentes. La colonización de las islas volcánicas, un tema que ya hemos tratado previamente, puede conducir a la radiación adaptativa. Un ejemplo son las numerosas especies de pinzones de Darwin en las islas Galápagos que descienden de una única especie de pinzón que colonizó originalmente el archipiélago y que a continuación se diversificó en numerosas especies que se especializaron en diferentes recursos alimentarios. A falta de otras aves en las islas, muchas “formas de vida”, lo que hoy llamaríamos “nichos”, estaban disponibles para que los ocuparan las distintas subpoblaciones de aves.
En resumen: la especiación empieza con las barreras al flujo de alelos
La separación geográfica completa, la separación geográfica parcial vista en las especies en anillo y el reparto de recursos entre subpoblaciones son todos barreras al flujo de alelos entre los miembros de lo que hasta entonces era una misma especie. Ello proporciona la oportunidad de que surjan alelos nuevos que no sean compartidos entre dos poblaciones y que se modifiquen las características medias de los dos grupos, alejándolos uno del otro. En el siguiente artículo de esta serie examinaremos algunos de los rasgos a los que contribuyen esos alelos; rasgos que promueven las barreras al flujo de alelos, promoviendo así la especiación.
Notas
- Nota del T.: En el original: “What goes around, comes around”


