14. De la Variación a la Especiación, Parte 4
31 de Mayo de 2013. Temas: Genética
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo examinamos los detalles de cómo el flujo de alelos queda prácticamente bloqueado entre dos poblaciones o especies que se han separado o divergido recientemente, al explotar diferentes nichos en una misma área geográfica.
En el artículo anterior, introducíamos la idea de que las especies se pueden formar en la misma localización geográfica mediante un reparto de los recursos, en el que las dos poblaciones se vuelven cada vez más adaptadas, a lo largo del tiempo, a la explotación de nichos diferentes. En el presente artículo vamos a examinar este fenómeno en detalle, utilizando un ejemplo de especie emergente que se ha formado en un pasado reciente y bajo la observación humana: la diversificación en las moscas de la manzana, Rhagoletis pomonella. Estas moscas son atraídas por los frutos aún inmaduros del espino albar, un pariente silvestre de los manzanos domésticos (es decir, algo que se parece a un pequeño manzano silvestre). El fruto del espino albar es también donde estas moscas encuentran pareja y ponen sus huevos, para que las larvas puedan alimentarse del fruto (y, de paso, hacen que se malogre y se caiga, con las larvas). Las moscas de la manzana producen sólo una generación al año, y sobreviven al invierno encerradas en fase de pupa. Además, tienen una corta vida adulta que les deja sólo un corto período de tiempo para encontrar pareja, aparearse, y para que las hembras pongan los huevos. Este período crucial, desde luego, está determinado por el ciclo vital del espino albar; cuándo su fruto está disponible para que las moscas lo utilicen como fuente de alimento y punto de encuentro. La selección natural, ejercida por el ciclo vital del espino, actúa como ta, sobre la variación genética relativa al momento de la eclosión en las poblaciones de moscas de la manzana. La fenología de la eclosión muestra una variación heredable y las moscas que eclosionan cerca de los momentos marginales cuando el fruto del espino está disponible (o peor, cuando no hay ningún fruto disponible) no se reproducen con tanto éxito como lo hacen las que eclosionan cuando los frutos del espino son abundantes. Por lo tanto, no en vano, el resultado es que observamos poblaciones de moscas de la manzana que están bien coordinadas en el tiempo con sus plantas hospedadoras, con la mayoría de los miembros de cualquier población de moscas de la manzana eclosionando de acuerdo con el máximo de disponibilidad de fruta:
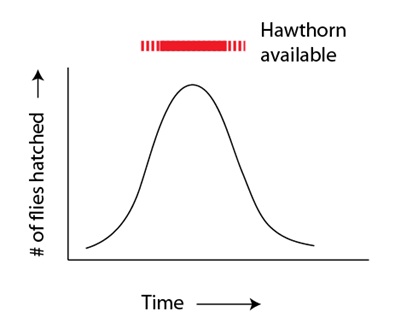
# de moscas eclosionadas
Disponibilidad de espinos
Tiempo
El momento de la eclosión es un ejemplo de un carácter o rasgo continuo; lo contrario que un carácter discontinuo. Los caracteres discontinuos son los que tienen distintas categorías: ojos negros frente a ojos azules, o flores rojas frente a flores blancas, y así sucesivamente. Muchos caracteres no pueden ser “clasificados” en tales categorías, sino que más bien forman una distribución en las poblaciones. Caracteres tales como la altura o el peso son ejemplos de caracteres continuos, y el momento de la eclosión de las moscas de la manzana es otro de ellos. El efecto que tiene el espino albar sobre las moscas de la manzana es un ejemplo de selección estabilizante. La disponibilidad de fruta selecciona negativamente las moscas que eclosionan fuera del momento óptimo, por uno u otro extremo; es decir, moscas que eclosionan demasiado pronto, o moscas que lo hacen demasiado tarde. El resultado global es que se mantiene la eclosión de las moscas sincronizada con la disponibilidad de fruta, generación tras generación.
Tentados por una manzana
Sin embargo, algo sucedió que alteró esta estable y equilibrada interacción: la introducción de las manzanas domésticas en Norteamérica por los colonos europeos. Como destacábamos antes, espinos albares y manzanos son plantas relacionadas, con frutos en cierto modo parecidos. Sin embargo, una diferencia era el momento del desarrollo del fruto en los manzanos en comparación con los espinos albares: los manzanos domésticos producen los frutos algunas semanas antes que los espinos. La introducción del manzano en el hábitat de las moscas proporcionó una fuente potencial de alimento a las moscas que eclosionaban en el extremo más “temprano” del espectro:
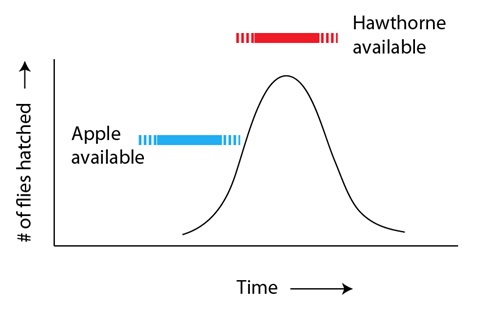
# de moscas eclosionadas
Disponibilidad de espinos
Disponibilidad de manzanas
Tiempo
Para las moscas “tempranas” que resultaron atraídas en su ambiente por este fruto nuevo, aunque similar, el resultado sería doble: (a) el descubrimiento de una nueva fuente de alimento con menos competencia por parte de los miembros de la propia especie, y (b) el encuentro de una pareja con esas mismas tendencias de preferencia por las manzanas. Lo que en principio era una combinación genética “perdedora”, (eclosionar demasiado pronto, sin comida suficiente ni perspectivas razonables de encontrar pareja), se había convertido ahora en la combinación “ganadora”. Como resultado. las variantes “tempranas” podían ahora reproducirse mucho más eficazmente que antes y, por tanto, aumentar su número en las generaciones siguientes:
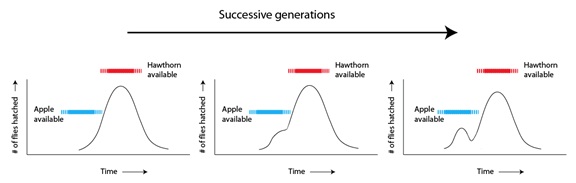
Generaciones sucesivas
Disponibilidad de espinos
Disponibilidad de manzanas
Tiempo
En otras palabras, una vez que aparecieron las manzanas, el ambiente dejó de seleccionar las poblaciones de moscas de forma estabilizadora, actuando más bien para moldear la variación en las dos subpoblaciones. La selección había cambiado ahora volviéndose una selección diversificadora. Es importante el hecho de que estas dos subpoblaciones no sólo se estaban diversificando con respecto al momento de eclosión y a la preferencia alimentaria, sino también, dada la naturaleza de su biología, con respecto a su preferencia de apareamiento. Cuando aumentaron las variedades de “manzana”, las moscas, de forma natural, empezaron a reproducirse con más frecuencia en esas otras variedades de “manzana” porque encontraban a sus parejas allí, en los manzanos. El resultado fue una barrera parcial al flujo de alelos que, con el tiempo, reforzaría las diferencias emergentes entre los dos grupos.
Aunque las “especies” de Rhagoletis pomonella del espino albar y del manzano han atraído el interés humano desde hace siglos, sobre todo debido al impacto económico de las mismas como una plaga, los genéticos están apenas empezando a hacerse una idea sobre las diferencias alélicas que eran los objetivos de la selección durante el proceso de separación. No sorprende pues, que los genes que afectan al momento de la eclosión, y que eran ya conocidos por la investigación anterior, demuestren ahora tener diferentes alelos en los dos grupos. Otros genes candidatos incluyen las proteínas receptoras que las moscas utilizan para detectar los olores de sus frutas preferidas (con ciertos alelos más afinados o ajustados con los olores de las manzanas, y otros afinados o ajustados con los olores de los frutos de los espinos albares). Lo que empezó siendo una variación dentro de una población se ha dividido ahora, por selección, en combinaciones alélicas adaptadas a distintos nichos y, dado el corto espacio de tiempo en que sucedió el cambio a las manzanas, es probable que las nuevas mutaciones no tuvieran un papel importante. Más bien, la recombinación y segregación de los alelos ya existentes de numerosos genes debió ser suficiente para producir las diferencias genéticas que volvieron más aptos a algunos miembros de la población original para aprovechar la nueva oportunidad. El efecto neto fue el cambio de unos pocos caracteres continuos (como el momento de eclosión y la preferencia por el olor de la fruta) para adaptarlos a un nuevo nicho ambiental, y provocar la aparición de una barrera al flujo de alelos.
Selección de las minorías
Una vez examinados los genes (y sus alelos) que estaban sometidos a la selección durante este proceso de especiación, podemos hacer algunas consideraciones más. El número de genes sometidos a la selección (y por lo tanto con diferentes alelos en las dos nuevas especies) será relativamente escaso. Sólo resultarán afectados los alelos que tienen que ver con rasgos o características relevantes para la adaptación al nuevo nicho. La mayoría de los genes seguirán siendo idénticos entre las dos poblaciones ya que no estaban sometidos a la selección diversificadora, sino que seguían bajo la selección estabilizadora por su papel (idéntico) en ambas especies. Por ejemplo, pensemos en los genes necesarios para la conversión de la energía en las células o para el desarrollo de las alas, procesos que ambas especies también necesitan realizar de igual manera. Estos genes tendrán los mismos alelos (o quizás sólo un alelo) en ambas poblaciones, porque la función de estos genes no era relevante para adaptarlas al nuevo nicho. Brevemente, el patrón general que la especiación produce será una pequeñísima muestra de diferencias entre los alelos de los genes sometidos a la selección o de los genes que por casualidad sufrieron la deriva, frente al conjunto de la amplia mayoría de genes que permanecieron idénticos porque no estaban sometidos a la selección, ni a la deriva.
En realidad, aparte del hecho de que los agricultores las vieron surgir, una de las razones por las que estamos seguros de que las moscas Rhagoletis pomonella “especializadas en” los espinos albares y en los manzanos, son realmente producto de una especiación reciente, es la abrumadora identidad entre sus genomas -- ellas sólo tienen diferencias mínimas en unos pocos genes. Biológicamente, sigue abierta la cuestión de si son realmente especies distintas porque siguen intercambiando alelos entre ellas aunque a una tasa muy reducida en comparación con el intercambio de alelos que realizan dentro de sus respectivas poblaciones. Como vimos con las especies en anillo, este ejemplo nos muestra que es posible observar, a día de hoy, las características precisas que podríamos predecir para un proceso de especiación “en marcha”. Además, demuestra que sólo un puñado de diferencias, derivadas de la variación ya existente dentro de la población, puede iniciar a dos subpoblaciones en una trayectoria que gradualmente aumente la barrera al flujo de alelos entre ellas. Con el tiempo, estos efectos pueden conducir a la formación de especies estrechamente relacionadas entre sí.
A largo plazo
La formación de las especies estrechamente relacionadas a partir de una población antecesora común es muy controvertida entre los cristianos evangélicos, aunque los mecanismos subyacentes no suelen ser entendidos. Sin embargo, lo más controvertido para muchos es la sugerencia de que estos mecanismos, en mayores lapsos de tiempo, pudieron también producir especies muy divergentes entre sí. En el siguiente artículo de esta serie volveremos sobre determinadas evidencias que apoyan la hipótesis de que especies modernas muy diversas, de hecho, derivan de poblaciones ancestrales comunes en un remoto pasado.
Lecturas complementarias
Schwarz, D. et al., (2009). Sympatric ecological speciation meets pyrosequencing: sampling the transcriptome of the apple maggot Rhagoletis pomonella. BMC Genomics 10; 633. (http://www.biomedcentral.com/1471-2164/10/633)


