33. El Ensamblado de los Planes de organización corporal de los Vertebrados, Parte 5
7 de Noviembre de 2013. Temas: Genética, Historia de la Vida
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.

Foto: Flickr user edenpictures [CC BY 2.0]
En este artículo discutimos el origen y diversificación de los mamíferos, incluida la evidencia genómica de la transición desde los huevos con vitelo hasta la nutrición del embrión a través de la placenta.
Como recordaremos del último artículo de esta serie, los tetrápodos del “grupo corona” son un grupo enormemente diverso, que va desde los anfibios a los reptiles, a las aves y a los mamíferos. Como este último grupo incluye a nuestra propia especie, no sería lógico desaprovechar esta oportunidad para discutir al menos unas cuantas características diferenciales entre los mamíferos y el resto de los tetrápodos, así como su historia evolutiva.
Mamíferos troncales: los sinápsidos
Los mamíferos son los únicos representantes vivientes del grupo de los llamados sinápsidos, un grupo que se separó del linaje dinosaurio/aviar (los saurópsidos), y que comenzaría a diversificarse a partir de finales del Carbonífero, hace unos 325 millones de años. Los numerosos sinápsidos que se conocen del registro fósil son mamíferos del grupo troncal, organismos relacionados con los mamíferos que se ramifican aparte del linaje que lleva hasta los mamíferos del “grupo corona”, que incluye a la última población ancestral de todos los mamíferos vivientes y a todas las especies descendientes de aquella población ancestral. El hecho de que parte de estos grupos troncales, los sinápsidos extintos, se conocieran hace tiempo como “reptiles mamiferoides”, y otros como “mamíferos reptilianos”, revela la naturaleza transitoria de sus caracteres que desdibujan la distinción entre “reptiles” y “mamíferos” en la forma que ahora ya conocemos: a través de la acumulación gradual de rasgos característicos del “grupo corona” de los mamíferos, y con un patrón de ramificación que indica el orden en que estos caracteres fueron adquiridos. Ejemplos de tales rasgos adquiridos incluyen la morfología de la mandíbula (la cooptación y reutilización, es decir la exaptación, de los huesos de la mandíbula para una nueva función auditiva especializada en el oído interno), el desarrollo del pelo y la lactancia (la secreción de leche para alimentar a las crías).
Del huevo a la placenta
A veces me encuentro a no biólogos, e incluso a algunos biólogos, que se sorprenden de que el “parto” no sea un carácter diferencial de los mamíferos o, más concretamente, de los mamíferos del “grupo corona”. La razón de esto estriba en que hay un linaje de mamíferos ovíparos, los monotremas, que aún tiene representantes vivientes. El hecho de que hoy en día existan mamíferos ovíparos significa que forman parte del “grupo corona” (por definición) y que, por lo tanto, cualquier carácter del que carezcan no podrá ser un carácter diagnóstico de dicho “grupo corona”:
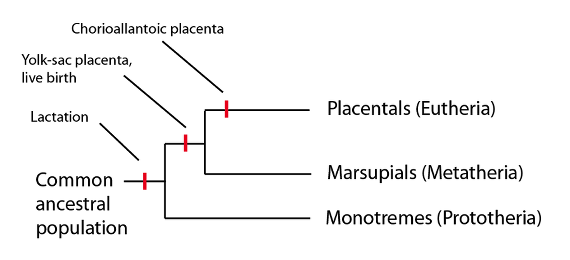
Los mamíferos del “grupo corona” incluyen tanto a los mamíferos ovíparos, los monotremas, como a los mamíferos no ovíparos, los marsupiales y los placentarios. De los caracteres mostrados en esta filogenia, sólo la lactancia es un carácter común a todo el grupo.
En los mamíferos ovíparos, como el ornitorrinco y las diversas especies de equidnas, las crías nacen de un huevo y luego son alimentadas con la leche de su madre, que es secretada en cierta zona de la piel y luego lamida por las crías. En los marsupiales, la gestación es mucho más corta que en los mamíferos placentarios y, tras el nacimiento, la cría, todavía muy embrionaria, se abre paso hasta una bolsa protectora en la que mamará de un pezón hasta completar su desarrollo. En los marsupiales se forma una breve conexión in utero entre la madre y el embrión, a través de la membrana del saco vitelino, para formar una placenta de “saco vitelino”. Sí, resulta algo complicado el que tanto marsupiales como mamíferos “placentarios”, es decir euterios, tengan ambos una placenta. La diferencia es que los mamíferos placentarios forman su placenta a partir de una membrana distinta: la membrana corioalantoidea.
Aunque monotremas y marsupiales no son mamíferos del “grupo troncal”, porque sus linajes persisten hasta la actualidad, podemos apreciar sus caracteres exactamente de la misma manera que lo hemos hecho con los grupos troncales. Dicho de otro modo, si los linajes de monotremas y marsupiales se hubieran extinguido, llamaríamos “mamíferos” a los euterios, y los monotremas y marsupiales serían grupos troncales en el linaje euterio. Al igual que hemos visto con los grupos troncales, los monotremas y los marsupiales demuestran que al estado euterio se llegó sólo a base de tiempo, y a través de una serie de pasos graduales. Aunque en realidad este proceso fue un gradiente, podemos arbitrariamente señalar algunas “etapas” en el camino:
- Puesta de huevos con lactancia tras la eclosión (estado monotrema)
- Gestación corta, con una placenta de saco vitelino con lactancia postnatal (estado marsupial)
- Gestación con una combinación de saco vitelino y placenta corioalantoidea (que prolonga el tiempo de gestación), con lactancia postnatal
- Reducción de la placenta de saco vitelino en favor de la placenta corioalantoidea (con tiempo de gestación aún más prolongado),con lactancia postnatal
- Gestación larga con placenta corioalantoidea, con lactancia postnatal (la condición euteria).
Por el camino, metaterios y euterios se irían apartando de la nutrición de sus embriones, in utero, basada en el vitelo, y pasarían a una nutrición basada en sus placentas. Al hacerlo, la maquinaria bioquímica de la nutrición basada en el contenido de vitelo del huevo iría siendo cada vez menos importante y, finalmente, del todo inútil. A nivel genético, los genes requeridos para la producción del vitelo dejarían de contribuir a la supervivencia o reproducción del organismo, lo que significa que dejarían de estar sometidos a la selección, y serían ahora libres de acumular mutaciones sin consecuencias para el organismo.
Buscando al muerto entre los vivos1
En el terreno práctico, cuando un gen deja de estar sometido a la selección, se mantiene ya sólo por la precisión general de la replicación del ADN durante la división celular. Aunque bastante precisa, la replicación del ADN no es perfecta. En las secuencias sometidas a la selección natural, las mutaciones son eliminadas de la población si los individuos que portan esas variantes no pueden reproducirse con la misma frecuencia que sus parientes no mutados. En los genes no sometidos ya a la selección natural, las mutaciones se acumularán lentamente con el tiempo.
En los mamíferos marsupiales y placentarios, uno de estos genes, llamado vitelogenina, era esperable que fuera liberado de la selección tras la aparición de la placenta. La vitelogenina es de importancia vital para la formación de la yema o vitelo del huevo, ya que, en los organismos ovíparos, actúa como principal portador de nutrientes desde el hígado hasta el huevo en formación. En 2008, un grupo de investigación estuvo buscando los residuos de las secuencias de la vitelogenina en los mamíferos monotremas, marsupiales y placentarios. Los monotremas, como era de esperar, tienen funcional la secuencia génica de la vitelogenina ya que son mamíferos ovíparos. Mientras que los mamíferos marsupiales y los placentarios carecen de secuencias génicas funcionales de la vitelogenina; pero tienen (muy) mutados los restos de las secuencias de la vitelogenina, indicando que estos linajes tuvieron un día la maquinaria bioquímica funcional para transferir nutrientes en masa a la yema del huevo. Esta observación encaja perfectamente a la luz de la predicción filogenética de que placentarios y marsupiales comparten una población ancestral común con los monotremas, y desde luego con los otros tetrápodos, siendo el oviparismo el estado ancestral, que luego se perdería en la población ancestral común de marsupiales y placentarios:
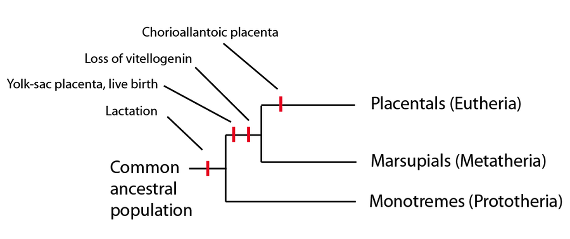
Así pues, la secuenciación del genoma nos permite verificar predicciones específicas sobre lo que deberíamos encontrar basándonos en filogenias construidas con caracteres anatómicos y morfológicos. En este caso, la presencia de las secuencias de la vitelogenina en los mamíferos placentarios, incluido el genoma humano, que nunca van a servir para formar el vitelo del huevo, es un notable ejemplo de confirmación de una predicción evolutiva; y es uno de los que siguen resultando muy problemáticos para los grupos antievolucionistas. En este contexto, sin embargo, la pérdida de la vitelogenina no fue sino un pequeño paso atrás en el camino hacia los linajes metaterios y euterios que, en contraste con los monotremas, siguen gozando de gran éxito a día de hoy.
En el siguiente artículo de esta serie exploraremos la diversificación de los mamíferos placentarios, incluido el linaje que conduce hasta nuestra propia especie: los primates.
Notas
-Nota del T.: En el original, “Seeking the dead among the living”


