40. Convertirse en Humanos, Parte 1: La Eva Mitocondrial y el Adán del Cromosoma Y
13 de Marzo de 2014. Temas: Adán, la caída y el pecado, Genética, Historia de la Vida, Origen del Hombre
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.

En este artículo situamos a la “Eva mitocondrial” y al “Adán del cromosoma Y” en su contexto paleontológico.
En el artículo precedente habíamos llegado, finalmente, a los orígenes de nuestra propia especie, Homo sapiens. Basándonos en los más antiguos restos conocidos en el registro fósil sabemos que los humanos modernos anatómicamente estaban ya presentes en África hace unos 200.000 años. Desde ese punto de partida, nuestra especie estaba lista para expandirse hacia Asia y Europa, empezando hace unos 100.000 años y de forma más significativa desde hace unos 50.000 años. Al hacerlo, seguiríamos, y luego nos encontraríamos, a las otras especies de Homo que habían salido de África antes que nosotros. Esos grupos incluyen a los Neandertales y a los Denisovanos, así como al Homo erectus que, como hemos visto, también salió de África y se distribuyó por toda Asia, incluidas las poblaciones de Indonesia que constituirían la base de los hallazgos pioneros de Eugêne Dubois. Neandertales y Denisovanos comparten una población ancestral común hace unos 400.000 años, aunque todavía no está claro si fue su población ancestral común la que salió de África, o si sus linajes primero se separaron en África y luego emigraron de allí ambos grupos independientemente. En cuanto al Homo erectus, los restos fósiles demuestran que tuvo una amplia distribución en Asia hace nada menos que 1.8 millones de años.
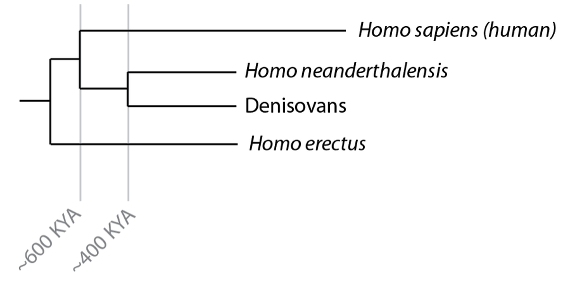
Relaciones de los homininosy momento aproximado de la divergencia de los linajes que conducen hasta los Neandertales, Denisovanos y humanos modernos.
Es alrededor de ese momento cuando llegamos al punto, en la evolución humana, del que muchos evangélicos al menos han oído hablar: el último antecesor femenino común de todos los humanos, conocido popularmente como la “Eva mitocondrial”, y de su equivalente masculino, nuestro último antecesor masculino común, popularmente conocido como el “Adán del cromosoma Y”.
La Eva mitocondrial y el Adán del cromosoma Y: antecesores comunes, pero no antecesores únicos
¡Espera un momento!, diremos: ¿Es que no hay pruebas suficientes de que la población humana desciende de una población que nunca fue inferior a los 10.000 individuos, aproximadamente, y de que es, por ello, un tema de una significativa consideración teológica? ¿Cómo es, entonces, que todos los humanos pueden compartir a una única mujer y a un único hombre como antecesores comunes? La respuesta breve es que todos los humanos sí comparten a un único hombre y a una única mujer como antecesores comunes, pero que ese hombre y esa mujer no son nuestros únicos o exclusivos antecesores. Más bien se trata de que ambos vienen de una población de unos 10.000 individuos, de cuya evidencia, y de las cuestiones teológicas que plantea, hablaremos en los siguientes artículos.
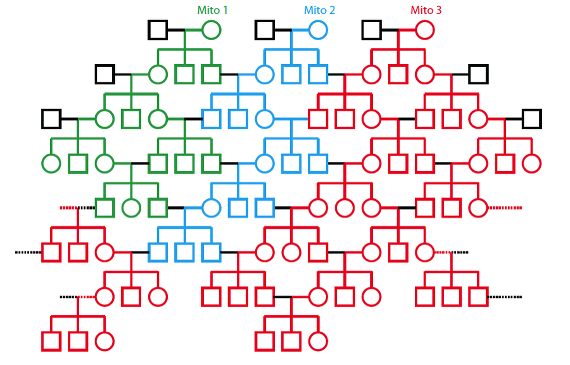
Para comprender cómo los humanos pueden tener un único antecesor maternal y paternal dentro de una población genéticamente diversa requiere por nuestra parte una corta excursión por la Genética y, específicamente, por las maneras en que se heredan ciertas formas de ADN. Como ya hemos tratado anteriormente, nuestras mitocondrias tienen su propio pequeño cromosoma como un recuerdo del tiempo en que fueron bacterias de vida libre. En la especie humana las mitocondrias se transfieren sólo de la madre al hijo: el esperma no aporta mitocondrias al huevo fecundado. Así que el ADN mitocondrial se hereda sólo a través del linaje materno, contrastando con el ADN cromosómico normal, que se hereda a través de ambos linajes, paterno y materno. El patrón maternal específico de herencia del ADN mitocondrial se presta el mismo a que ciertas variantes mitocondriales “invadan” una población, cosa que podemos ilustrar mediante un gran árbol familiar, un linaje o un pedigrí. (Nota sobre los símbolos del pedigrí: los círculos representan hembras; los cuadrados, machos; una barra horizontal uniéndolos representa un emparejamiento, y una barra vertical desde un emparejamiento conecta con los hijos o descendientes de ese emparejamiento.)
En el linaje o pedigrí de abajo vemos una gran familia que muestra la herencia de tres variantes mitocondriales (etiquetadas con diferentes colores). Para mantener el linaje lo bastante compacto como para poder mostrarlo, las líneas discontinuas indican emparejamientos que conectan unos con otros de un lado y del otro de forma envolvente. Como podemos ver, la variante roja “Mito 3” ha “invadido” esta población. Todos los individuos de las generaciones más recientes de esta familia comparten a la mujer de la generación de arriba a la derecha como antecesora común de sus mitocondrias:
Con este mismo pedigrí, sigamos la pista ahora algunas variantes hipotéticas del cromosoma Y. Los cromosomas Y se transfieren, obviamente, sólo de padres a hijos, dado un patrón de herencia específicamente paterno. Este patrón, como el patrón específicamente maternal de la herencia mitocondrial, puede también conducir fácilmente a la invasión de la población por parte de ciertas variantes. Supongamos que esta misma familia tiene también tres variantes del cromosoma Y en las generaciones más antiguas:
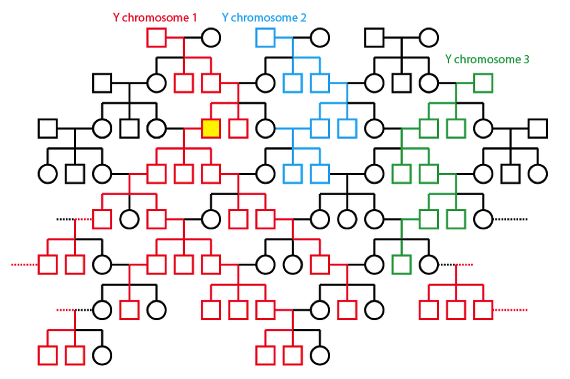
En este caso, la variante del “cromosoma Y - 1” invade la población, y todos los individuos en las generaciones más recientes tienen al hombre sombreado en amarillo como su más reciente antecesor masculino en común.
Ahora que ya hemos identificado una hembra y un macho como antecesores comunes de las generaciones más recientes en este pedigrí, podemos ilustrar que ellos no son sus únicos antecesores. Tanto la Eva “mitocondrial” como el “Adán” del cromosoma Y de esta familia proceden de una población mayor; y podemos fácilmente mostrarlo viendo la variación presente en el ADN cromosómico regular, el que se transfiere tanto a través de los linajes maternales como paternales.
Volvamos otra vez al mismo pedigrí anterior, pero esta vez ilustremos la variación en el ADN cromosómico regular con diferentes colores. Ahora resulta mucho más difícil para esta variación invadir la población a corto plazo, con rapidez, porque esta variación puede transferirse tanto por los machos como por las hembras:
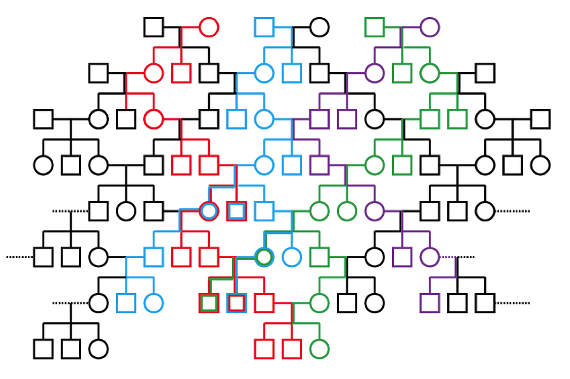
En contraste con los patrones del ADN mitocondrial y del ADN del cromosoma Y, vemos una diversidad de variación en el ADN cromosómico regular transmitida desde las primeras generaciones hasta las más recientes. Por ejemplo, consideremos la pareja central de la primera generación. Mientras su variación mitocondrial y cromosómica-Y se ha perdido en esta población, la variación cromosómica normal del macho (representado en la línea azul), ha llegado hasta el momento actual sin ningún problema. Y por lo tanto tenemos un “registro” de su genealogía en la población, incluso tras la pérdida de su variación del cromosoma Y. De igual manera, consideremos la hembra de la pareja de la izquierda en la primera generación. Aunque su variación mitocondrial se ha perdido, su variación cromosómica normal (representada por la línea roja), sí se ha ido transmitiendo. La cantidad total de variación genética en los cromosomas normales es, por tanto, una herramienta para determinar cuántos antecesores tiene esta población.
Es esta variación en el ADN cromosómico normal la que indica que esta población no ha sufrido una reducción drástica de su tamaño poblacional en el pasado reciente; y que, incluso pudiendo señalar los antecesores comunes recientes del ADN mitocondrial y de los cromosomas Y, estos antecesores comunes proceden de una población genéticamente diversa. Lo mismo sucede con nuestro propio linaje: nosotros tenemos también tanto un antecesor maternal común de nuestro ADN mitocondrial, la “Eva mitocondrial”, como un antecesor paternal común de nuestro ADN del cromosoma Y, el “Adán cromosómico”. Sin embargo, la diversidad de nuestro ADN cromosómico normal nos muestra que esos individuos eran parte de una población amplia y genéticamente diversa. Como en el ejemplo con el que hemos trabajado, sabemos todo esto por la diversidad del ADN cromosómico normal que vemos en las poblaciones humanas modernas.
Así que, ¿A qué se debe tanto entusiasmo por estos dos individuos? En muchos aspectos esto ha sido exagerado. El único mérito de estos dos individuos es el ser los últimos antecesores comunes de sólo una pequeña parte de nuestros genomas (respectivamente del ADN mitocondrial y del ADN del cromosoma Y). Mientras tanto, hay un hecho interesante: esos dos individuos no eran sensiblemente diferentes de otros de sus respectivas poblaciones. Si los científicos no les hubieran etiquetado con nombres aludiendo a la narrativa bíblica, probablemente hubieran sido poco conocidos entre los cristianos.
Situando a la Eva Mitocondrial y al Adán del cromosoma Y en el tiempo
Las estimaciones actuales sitúan a la Eva mitocondrial inmediatamente después de los albores del Homo sapiens, tal como aparece en el registro fósil, hace unos 180.000 años. Esto la coloca dentro de nuestra especie. Hasta hace poco, el Adán del cromosoma Y se databa con posterioridad, hace unos 50.000 años, el momento de la migración humana significativa fuera de África. Recientemente, sin embargo, se ha encontrado una rara variante del cromosoma Y en humanos modernos que retrasa el último antecesor común de todos los ADN cromosómicos humanos hasta hace 210.000 años, lo que de forma muy interesante, lo sitúa precisamente en el umbral de nuestra propia especie, según el registro fósil. Como nuestra especie surgió como una población continua que divergió gradualmente de otros homininos, no hay razón alguna para esperar que toda la variación de nuestro ADN proceda de un antecesor común (que sea coalescente, por utilizar el término técnico) dentro de nuestra especie. De hecho, parte de nuestra variación cromosómica normal no es coalescente dentro de nuestra especie, ni siquiera remontándonos tan atrás en el tiempo como a nuestra población antecesora común con los chimpancés. Como ya hemos discutido antes, “especie” es un término de conveniencia que los biólogos utilizan para trazar una línea en lo que en realidad es un gradiente de cambio gradual; y, biológicamente, nuestra especie no es una excepción.
En el siguiente artículo vamos a seguir explorando las borrosas fronteras de nuestra propia especie a medida que viajamos con algunos de nuestros antecesores hacia fuera de África, y encontramos a otras especies de homininos en el proceso.
Lecturas complementarias:
- Tim Keller on the historical view of Adam
- Dennis Alexander on the representative and symbolic views of Adam
- “Why the Church Needs Multiple Theories of Original Sin”
- James Kidder: “The Human Fossil Record”


