35. La Revolución Placentaria, Parte 2
12 de Diciembre de 2013. Temas: Genética, Historia de la Vida
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.

Foto: Dallas Krentzel (Creative Commons Attribution 2.0 Generic)
En este artículo tratamos sobre el primer primate conocido y sobre un suceso fortuito que lastró a algunos grupos de primates (incluida nuestra propia especie) con la incapacidad de sintetizar su propia vitamina C.
En el último artículo de esta serie examinábamos la rápida diversificación de los mamíferos placentarios tras la extinción de los dinosaurios no aviares (provocada por el impacto del asteroide que marcó el límite entre el Cretácico y el Paleógeno, hace 66 millones de años). No mucho más tarde, en términos geológicos, los primates aparecen en el registro fósil. Como nuestra propia especie está anidada dentro de los primates, cosa reconocida por primera vez por Linneo en los años 1700, el origen de este grupo ha tenido mucho interés para los paleontólogos.
Actualmente el primate más antiguo conocido es un animalillo procedente de mediados del Paleógeno de lo que hoy es la China actual. Este diminuto primate, Archicebus achilles, es un haplorrino del grupo corona que vivió hace unos 55 millones de años. Los haplorrinos son un grupo monofilético que incluye a los actuales tarseros y a los antropoides, o especies de “aspecto humano”: los monos del Nuevo y del Viejo Mundo y los grandes simios, incluida la especie humana. Los tarseros y antropoides están más estrechamente emparentados entre sí que con los lemures o lémures, otro grupo de primates con representantes actuales:
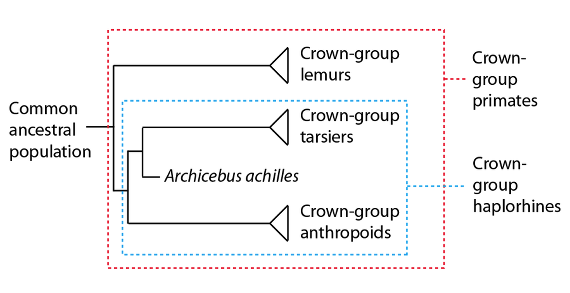
Relaciones filogenéticas entre los primates del “grupo corona”, mostrando la posición de Archicebus achilles como un tarsero del “grupo troncal”, cercano a la base del árbol de los primates.
Más allá de su edad y su estatus actual como primate más antiguo conocido, Archicebus achilles resulta también notable por estar situado, al parecer, muy cerca de la última población ancestral de los linajes del “grupo corona” de los tarseros y del “grupo corona” de los antropoides; aunque parece más estrechamente emparentado con el linaje de los tarseros que con el linaje de los antropoides (es decir, a pesar de estar muy cerca del último antecesor común de tarseros y antropoides, Archicebus se interpreta, más bien, como un tarsero del “grupo troncal”). Los caracteres de Archicebus, y su posición cerca de la base del linaje de los primates, sugieren que la última población ancestral común de los primates era también de pequeña estatura, arborícola y que se alimentaba de insectos. La posición de Archicebus apoya también la hipótesis de que los primates evolucionaron primero en Asia, emigrando más tarde a África (donde, como veremos más adelante en otro artículo, evolucionaría el linaje que conduce hasta los seres humanos).
Adiós a GULO
En la base del árbol de los haplorrinos es donde tuvo lugar un suceso que ya hemos considerado previamente como una firme evidencia de la existencia de un antecesor común para el ser humano y para los grandes simios; un suceso que a muchos cristianos, que ya han revisado las pruebas de la evolución, les resulta familiar. Una característica que comparten los primates haplorrinos es su curiosa incapacidad para sintetizar su propia vitamina C, que les obliga a obtenerla a través de la dieta. Esto no es lo normal: la mayoría de los mamíferos, incluidos los primates no haplorrinos, son capaces de sintetizar vitamina C a partir del azúcar básico de su dieta, la glucosa. La razón genética de esta deficiencia se ha averiguado: en los primates haplorrinos falta la enzima responsable del último paso en la biosíntesis de la vitamina C (llamada L-gulonolactona oxidasa; abreviada como “GULO”). Sin embargo, y a pesar de esta carencia de función enzimática, los restos mutados del gen que codifica para la enzima GULO sí se encuentran en esos primates. Como esta deficiencia afecta a todos los primates haplorrinos, la explicación más sencilla es que esta función enzimática se perdió una única vez, antes de la divergencia del “grupo corona” de los haplorrinos:
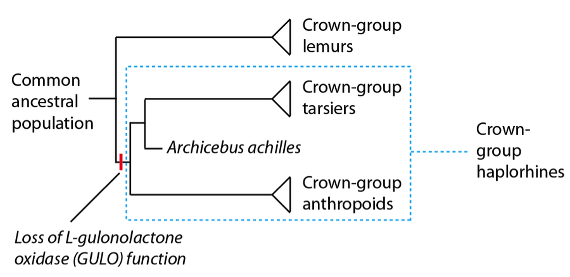
Para apreciar los detalles moleculares de la pérdida de GULO en el linaje de los haplorrinos, vamos a revisar brevemente la estructura y función del gen, cosa que ya hemos tratado anteriormente con más detalle:
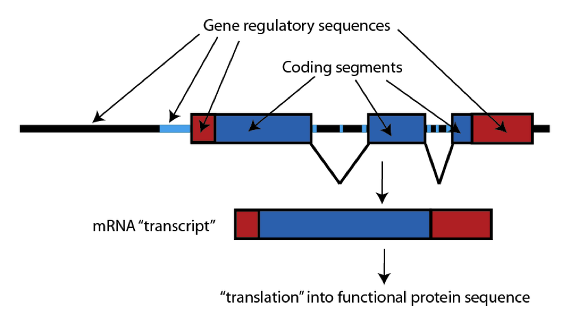
Los genes tienen una estructura típica (obviamente algo simplificada aquí). En primer lugar, está la secuencia de ADN actual que especifica la secuencia del producto proteínico (la llamada “secuencia codificante”, mostrada en azul). Esta secuencia, generalmente está dividida en segmentos en los genes de los mamíferos, y esas secuencias son empalmadas cuando la secuencia de ADN del gen se transcribe a una “copia de trabajo” llamada ARNm, un duplicado abreviado del código que puede utilizarse por la maquinaria celular para construir realmente la proteína especificada.
El segmento de un gen que se empalma en un ARNm se conoce técnicamente como “exón”; y los genes GULO funcionales en mamíferos tienen doce exones. Al comparar la secuencia del gen GULO en los grandes simios (homínidos), que son los haplorrinos del “grupo corona”, vemos que muchos de los exones del gen GULO se han suprimido por completo, eliminando totalmente la posibilidad de que sea funcional. En la especie humana, por ejemplo, sólo quedan los exones 4, 7, 9, 10 y 12; y los chimpancés y macacos tienen el mismo patrón:
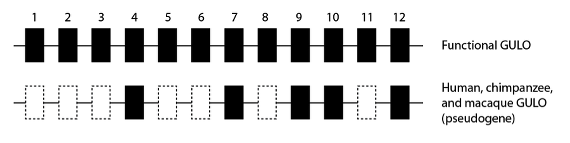
Diagrama esquemático de un gen GULO funcional comparado con el estado observado en varios haplorrinos del “grupo corona” de los que se dispone de datos sobre la secuencia del genoma. Los mismos siete exones en seres humanos, chimpancés y macacos se han eliminado del seudogen GULO, eliminando la función de la enzima.
No es probable que todas las deleciones que eliminaron estos exones fueran las mutaciones originales que suprimieron la función de este gen en el linaje de los haplorrinos. Más bien, estas deleciones tuvieron lugar probablemente después de la primitiva pérdida de la función del GULO, cuando las nuevas mutaciones en esta secuencia, ahora ya no funcional, dejarían de tener consecuencias sobre un gen que era ya defectuoso. Sin embargo, las deleciones que vemos compartidas en los grandes simios (homínidos) ocurrieron antes de que estas especies tomaran sus caminos separados, y fueron heredadas por la especie resultante de este grupo, exactamente de la misma manera que hemos discutido ya anteriormente en esta serie, cuando dedicamos varios artículos estableciendo la analogía entre genomas y “textos copiados” que con el tiempo van acumulando errores.
Mutaciones compartidas: un desafío permanente a los puntos de vista antievolutivos
Aunque observar genes defectuosos con mutaciones idénticas en grupos de especies anidadas es fácil de entender desde un punto de vista evolutivo, esta cuestión sigue siendo un problema extremadamente difícil de explicar para las organizaciones antievolucionistas. De hecho, el único recurso es tratar de argumentar que las mutaciones idénticas que observamos en las numerosas especies no fueron heredadas de un antecesor común, por lo que debieron haber sucedido independientemente en cada una de esas especies. La probabilidad de que mutaciones idénticas se den numerosas veces en especies independientes es, desde luego, extremadamente pequeña; por no mencionar que tales sucesos acaben produciendo precisamente el patrón filogenético que predicen todas las demás evidencias.
Así que, a pesar de la desafortunada pérdida del GULO, con el consiguiente requisito de obtener vitamina C con la dieta, el linaje antropoide continuaría su expansión y diversificación. En el siguiente artículo de esta serie vamos a seguir examinando la diversificación de este grupo y la aparición de nuestro propio grupo dentro de él: el género Homo.
Lecturas complementarias:
- Lachappelle, M.Y. and Drouin, G. (2011). Inactivation dates of the human and guinea pig vitamin C genes. Genetica 139: 199-207. http://www.ncbi.nlm.nih.gov/pubmed/21140195


