39. Del Primate al Humano, Parte 4
27 de Febrero de 2014. Temas: Genética, Historia de la Vida, Origen del Hombre
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.

En este artículo analizamos la situación actual de la Paleontología de los “homininos” y trataremos de situar en ella a nuestro linaje.
En el artículo precedente dejábamos la historia de la Paleontología de los homininos a principios de los años 1950, con el hombre de Piltdown desenmascarado, y con las correctas interpretaciones del Australopithecus africanus de Dart y del Pithecanthropus (Homo) erectus de Dubois siendo ya aceptadas por la comunidad científica. Los paleontólogos sabían ahora dónde buscar: en África, como Darwin sugirió. Y también sabían qué buscar: una especie con cuerpo de aspecto humano y capacidad craneal subhumana. En las décadas siguientes aparecerían muchos más hallazgos, revelando a los homininos como un gran grupo, con numerosas especies. Más que encontrar una “escalera” de especies que condujera a la especie humana, la Paleontología de los homininos descubriría un árbol ramificado con nuestros parientes cercanos.
Como pasa con todos los grupos de especies fósiles, resulta estimulante determinar las relaciones precisas entre ellos. Y como ya hemos señalado con anterioridad, la línea que delimita las “especies” es siempre borrosa, imprecisa y su separación se logra sólo lentamente y con el tiempo. Las especies muy distantes son fáciles de identificar como diferentes pero, a medida que se recogen más y más datos, las “lagunas” existentes entre ellas empiezan a rellenarse, haciendo más difíciles las distinciones. En una disciplina como es la Paleontología de los homininos este problema se agudiza aún más debido al gran interés que suscita nuestro propio linaje, lo que significa que hemos invertido un gran esfuerzo para encontrar tantos restos de homininos como nos ha sido posible. Con tantos datos el cuadro global se vuelve cada vez más difícil de resolver en detalle, a pesar de que las líneas generales estén cada vez más claras. Esto determina que algunos paleontólogos tiendan a describir un gran número de especies de homininos fósiles, mientras que otros prefieren agrupar los restos de los homininos en un menor número de especies. De manera informal, a los que prefieren el mayor número de especies se les llama “separatistas” o “segregadores” (“splitters”), mientras que a los que prefieren menos especies se les llama “unionistas” o “aglutinadores” (“lumpers”). El hecho de que tal debate sea siquiera posible, es una prueba del sólido conjunto de datos de carácter gradual de los que disponemos.
Paleontología de los homininos: un esquema conciso de las pruebas actuales
En líneas generales, el estado actual de la Paleontología de los “homininos” reconoce varias grandes agrupaciones, o grados, con límites nada claros, como cabía esperar. A continuación mostramos una filogenia de consenso que representa el pensamiento actual, que nos presenta al grupo de los homininos como el árbol denso que es en realidad y destacando la abundancia de pruebas acumuladas desde los tiempos de Dart y Broom:
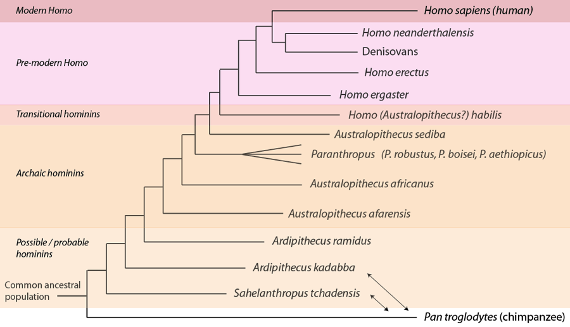
Filogenia de los homininos basada en pruebas paleontológicas actuales, y en algún caso genómicas, Disponemos de secuencias genómicas de humanos, denisovanos y neandertales. Como ya hemos comentado previamente, Sahelanthropus tchadensis y Ardipithecus kadabba pueden no ser homininos: podrían estar menos estrechamente relacionados con los humanos que con los chimpancés, en cuyo caso sus posiciones tendrían que ser intercambiadas con las de los chimpancés en la filogenia (indicadas con flechas de dos puntas). Varias de las especies de homininos no se representan. Ver detalles en el texto.
Avanzando desde el pasado hacia el presente, el primer grupo que encontramos incluye a las especies que hemos comentado antes, especies que se sitúan cerca de la última población ancestral común de humanos y chimpancés. Este grupo, conocido como de “homininos posibles y probables”, incluye a las especies de “posibles” homininos: Sahelanthropus tchadensis, Orrorin tugenensis, y Ardipithecus kadabba; pero sólo a un hominino “probable”: Ardipithecus ramidus. Como señalamos antes, la evidencia de bipedestación es más fuerte en Ardipithecus ramidus. Mientras que los “separatistas” o “segregadores” reconocen a estas especies como diferentes, los “unionistas” o “aglutinadores” las agrupan todas juntas como Ardipithecus ramidus sensu lato; donde “sensu lato” significa “en sentido amplio”. En general este grupo exhibe un pequeño volumen de caja craneana (menos de 350 cm3) y ciertos indicios de bipedalismo.
El segundo grupo, el de los “homininos arcaicos”, incluye a australopitecinos tales como el Australopithecus africanus de Dart y una especie anterior, el Australopithecus afarensis, el más famoso de los cuales es el apodado “Lucy”. También dentro de este grupo general están los homininos arcaicos “robustos”, las diversas especies clasificadas como Paranthropus (tales como el Paranthropus robustus de Bloom). Por lo general, en este grupo vemos una evidencia creciente de bipedalismo ocasional (es decir, facultativo), y un nuevo incremento del volumen craneal hasta el rango de los 500 cm3. Aunque los” separatistas o segregadores” ven numerosas especies más en este grupo, los “unionistas o aglutinadores” reconocen sólo a A. afarensis, A. africanus, y a una especie que incluye a todos los Paranthropus.
El tercer grupo, los “homininos de transición” incluye a los miembros más antiguos que se conocen de Homo, tales como por ejemplo el Homo habilis. Sin embargo, para subrayar su naturaleza transicional, algunos autores no sitúan a esta especie dentro del género Homo sino, más bien, lo describen como un australopitecino, Australopithecus habilis. La capacidad craneana de habilis va desde los 500 a los 700 cm3, aproximadamente. Otra especie parecida, Homo rudolfensis, es reconocida como tal por los separatistas o segregadores, pero los unionistas o aglutinadores las agrupan juntas como H. habilis sensu lato. Estas especies, (o esta especie según consideremos), muestran una creciente evidencia del bipedalismo, pero no un compromiso total con la forma de vida bípeda (es decir con el bipedalismo obligado) que veremos, más tarde, en los Homo posteriores. Estos grupos utilizaban también herramientas de piedra del tipo Oldovayense, una tecnología de herramientas de piedra relativamente sencilla, llamada así por el sitio de su primer hallazgo.
El cuarto grupo, el de los “Homo premodernos” incluye al Homo ergaster y al Homo erectus de Dubois, las primeras especies de Homo que fueron por completo bípedas. Aunque los separatistas o segregadores reconocen a H. ergaster y H. erectus como especies distintas, los unionistas o aglutinadores las reúnen como Homo erectus sensu lato. Este grado incluye también a los Neandertales más familiares así como a los Denisovanos, especies de las que tenemos secuencias genómicas completas. La capacidad craneana en este grado va desde los 700 cm3 de los primeros Homo ergaster / erectus, hasta el volumen que encontramos en los seres humanos modernos, de unos 1400 cm3 (y, de hecho, supera el volumen del humano medio moderno en el linaje neandertal, en el que se han llegado a encontrar cráneos con 1600 cm3). Al principio, en este grado, se observa ya el avance hacia el desarrollo de la tecnología de herramientas de piedra más compleja del Achelense.
El último grupo, el del “Homo moderno”, contiene sólo a nuestra propia especie, Homo sapiens, que aparece en el registro fósil hace unos 200.000 años. Los separatistas o segregadores, sin embargo, han propuesto una subespecie temprana, Homo sapiens idaltu, hace unos 160.000 años.
Antecesores, puede; parientes, seguro.
Esta diversidad de restos, por una parte complica los detalles de nuestra evolución, porque no podemos estar del todo seguros de cuál de estos grupos es el antecesor directo de nuestra propia especie, si es que lo es alguno; y por otra, clarifica el cuadro general, porque éstos son nuestros parientes cercanos incluso aunque no fueran nuestros antecesores directos. En conjunto, podemos estar seguros de que nuestro linaje pasó por todos esos grados sucesivamente: como ardipitecino (o un pariente cercano), hacia un australopitecino (o un pariente cercano), y así sucesivamente a través de los homininos de transición y hasta los Homo premodernos, y luego los modernos. Por el camino, nuestro linaje aumentaría su capacidad craneana media, adquiriría el uso de herramientas cada vez más sofisticadas y, hace unos 200.000 años, tomaría nuestra forma actual en África, antes de nuestra expansión por todo la Tierra.
En el artículo siguiente retomaremos la historia en el momento de la expansión de nuestra especie fuera de África, y de su encuentro con los descendientes de otros linajes relacionados que habían salido de África antes que nosotros.
Lecturas complementarias:
- Wood, B. and Lonergan, N. (2008). “The hominin fossil record: taxa, grades and clades.” Journal of Anatomy 212: 354 – 376.
- James Kidder: “The Human Fossil Record.”


