23. Evolución Convergente y Homología Subyacente
15 de Agosto de 2013. Temas: Genética
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo examinamos cómo la evolución convergente resulta favorecida por homologías subyacentes.
En el último artículo de esta serie introdujimos el concepto de homoplasia, una similitud de forma en dos linajes que surge debida a sucesos independientes. En el ejemplo que veíamos entonces, las aves y los murciélagos independientemente habían logrado volar activamente a través de una evolución convergente, con la que los murciélagos llegaron a alas basadas en una membrana y las aves a alas cubiertas de plumas. Como la última población ancestral común de aves y murciélagos no era capaz de volar activamente, éste es un buen ejemplo de homoplasia surgida por evolución convergente.
Sin embargo, en el trasfondo de esta convergencia hay una conexión más profunda. Los murciélagos y las aves son tetrápodos ambos, organismos con espina dorsal y cuatro extremidades. El plan de organización corporal de los tetrápodos era ya una característica de su población antecesora común más reciente y se ha mantenido en ambos linajes. Por lo tanto, cuando se consideran estrictamente como miembros anteriores, las alas de murciélagos y aves son estructuras homólogas. Tanto en aves como en murciélagos, los miembros anteriores han sido moldeados para el vuelo, a través de la selección natural, de diferente manera; pero el punto de partida de ambas alas era una estructura homóloga. En otras palabras, subyacente al fenómeno de la convergencia en el vuelo activo de murciélagos y aves persiste una homología más profunda - la extremidad a partir de la cual ambos linajes construyen independientemente sus alas. Para representar esto en una filogenia, deberíamos situar el plan de organización corporal de los tetrápodos anteriormente a la divergencia de todos ellos y el vuelo activo como dos sucesos independientes en sus linajes correspondientes:
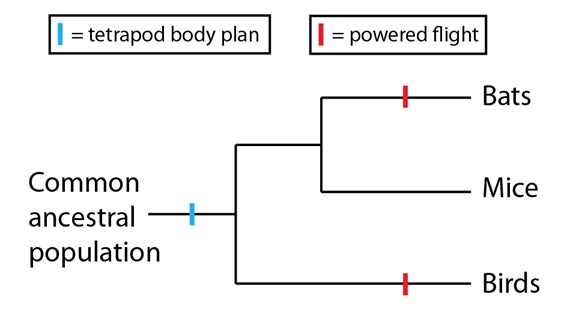
Este patrón (sucesos convergentes con homologías latentes subyacentes más profundas) es uno de los que se ven repetidas veces en la evolución. De hecho, estas homologías más profundas mejoran las probabilidades con la que sucederán los procesos convergentes porque proporcionan una base común que linajes diferentes pueden emplear para una innovación independiente. Para los murciélagos y las aves, las adaptaciones que conducen al vuelo fueron posibles porque ambos linajes poseían miembros anteriores susceptibles de ser modificados, con el tiempo, de una función a otra. Aunque este ejemplo se sitúa a nivel anatómico, este tipo de “predisposiciones” y los procesos convergentes que surgen de ellos pueden verse también a nivel molecular.
Los ojos lo tienen1
Como mencionábamos en un artículo anterior de esta serie, los ojos en cámara son uno de los ejemplos más llamativos de evolución convergente y han aparecido independientemente en varios linajes; los ejemplos más conocidos de ellos son los vertebrados, los cefalópodos como pulpos y calamares, y ciertas medusas. Los ojos en cámara tienen un estrato de células fotosensibles, la retina, así como una lente que enfoca la luz sobre la retina. Explicar la distribución de los ojos en cámara en estos tres grupos nos obliga a recurrir a tres sucesos convergentes en su filogenia (las medusas forman parte de los Cnidarios):
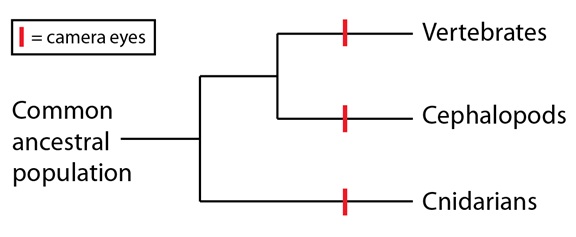
A primera vista, parece altamente improbable que tres linajes tan lejanamente relacionados fueran a convergir en una estructura tan notable como el ojo en cámara. Al final, sin embargo, una homología clave entre los tres grupos resuelve en gran medida esas dificultades: las moléculas que funcionan como sensores de luz.
En su forma más básica, la percepción del medio ambiente exterior requiere que dicho ambiente induzca un cambio en las células. En consecuencia la percepción de la luz requiere un cambio de algún tipo, inducido por la propia luz. Las moléculas clave que realizan esta función en los tres grupos anteriormente mencionados son las proteínas llamadas opsinas y sus asociados químicos, un grupo de compuestos llamados retinaldehidos o Vitamina A-aldehidos. Cada proteína opsina tiene un retinaldehido unido a ella, y juntos, la pareja opsina/retinaldehido, actúan como un sensor de luz. Los retinaldehidos cambian de forma al interaccionar con la luz (es decir, absorben un fotón, representado por la letra gamma en el esquema siguiente). Este cambio de forma, a su vez, altera la forma de la proteína opsina unida al retinaldehido:
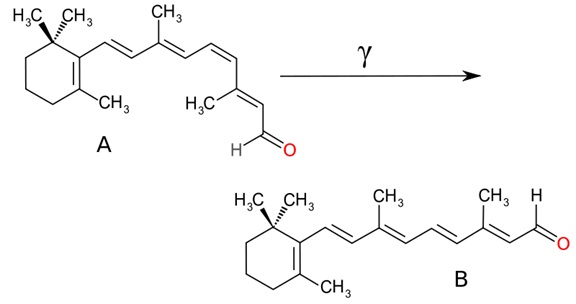
Fuente: http://en.wikipedia.org/wiki/ Archivo:RetinalCisandTrans.svg
El cambio de forma de la opsina afecta al flujo de carga eléctrica en las células responsables de la percepción de la luz, y estos cambios en la carga eléctrica son los que se perciben e interpretan como “luz” por el cerebro.
El sistema opsina/retinaldehido para la detección de la luz está muy generalizado: de hecho, todos los animales que pueden detectar la luz utilizan estas moléculas como sustrato físico para hacerlo, tengan ojos en cámara u otros tipos de ojos, tales como los ojos compuestos, o las simples agrupaciones de células fotosensibles. Esto supone una fuerte evidencia de que el sistema opsina/retinaldehido precede a la divergencia de los tres grupos que hemos considerado aquí:
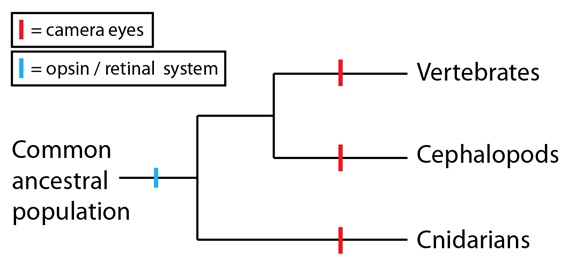
Con esta información en mente, podemos ver que el desarrollo de los ojos en cámara en estos linajes no es tan improbable como podríamos haber pensado al principio. En los tres casos, estos linajes forman un ojo en cámara en torno a un sistema molecular de detección de luz preexistente. Los ojos en cámara, en sí mismos, podrían ser convergentes pero están basados en una homología subyacente más profunda, que resuelve las dificultades que pudieran aparecer de plantearse a través de sucesivas modificaciones de un sistema ancestral. Y como vimos en el caso de las alas de murciélagos y aves, también hay diferencias entre los ojos en cámara en esos linajes lo que apoya la hipótesis de que son el resultado de una evolución convergente; el ejemplo mejor conocido de dichas diferencias es el hecho de que, en los ojos de vertebrados y cefalópodos, el “cableado” de nervios está dispuesto en direcciones opuestas.
Oír es creer2
Otro ejemplo de “predisposición molecular” que lleva a la convergencia puede verse en la maquinaria molecular subyacente en otra forma de percepción sensorial: la audición de ultrasonidos requerida por la ecolocación en los murciélagos y en odontocetos3. Ambos grupos utilizan la ecolocación para la navegación y para localizar a sus presas en ambientes en los que la percepción visual está muy limitada o incluso es del todo imposible. La evidencia de que el desarrollo de la ecolocación se debe a una convergencia, en estos dos grupos de mamíferos tan divergentes, es muy grande: ningún otro grupo de mamíferos, estrechamente relacionado con alguno de ellos, ha desarrollado esa capacidad.
La base celular / molecular de la detección del sonido en los mamíferos está en un conjunto de células en el oído que extienden unas prolongaciones que parecen pelos, llamadas cilios, que vibran en respuesta a las diferentes longitudes de onda del sonido. Los cilios también cambian su longitud y sus propiedades vibratorias en respuesta a distintos estímulos auditivos. Las vibraciones se utilizan para cambiar el flujo de carga eléctrica en esas células, que lleva al final señales al sistema nervioso que el cerebro interpreta como sonidos. Todos los mamíferos utilizan una proteína llamada prestina como parte de su sistema auditivo. La prestina es una “proteína motriz” que puede cambiar la forma de la célula desplazando sus estructuras internas y que los mamíferos la utilizan para modificar los cilios en respuesta al sonido.
Sabemos que el sistema cilios/prestina es anterior a los mamíferos, así que no es de extrañar que los odontocetos (como delfines y orcas) y los murciélagos lo utilicen para oír. Lo que resulta interesante es, sin embargo, que la proteína prestina se haya moldeado independientemente, a través de selección natural, para sintonizar con el sonido de elevada frecuencia (es decir, ultrasónico) más útil para la ecolocación. De hecho, en una filogenia restringida a las secuencias de la prestina, las prestinas de murciélago y de odontoceto aparecen como las más estrechamente relacionadas entre sí; un hecho en clara contradicción con el árbol filogenético de especies de murciélagos y ballenas. Sin embargo, un examen más detallado muestra que estas sorprendentes similitudes son el resultado de una evolución convergente y que no son debidas a que se comparta un antecesor común más reciente. En ambos casos, la proteína prestina estaba disponible para ser sintonizada con las ondas de sonido ultrasónicas y mutaciones similares, aunque no idénticas, fueron seleccionadas en ambos linajes sobre la marcha (otro ejemplo de una “homología subyacente” favoreciendo sucesos convergentes independientes)
Resumiendo: la evolución como proceso no aleatorio
Un error muy común sobre la evolución es el pensar que se trata de un proceso predominantemente aleatorio (que está influido sobre todo por sucesos aleatorios). Aunque ya hemos mostrado que la evolución tiene un fuerte componente no aleatorio (la selección natural), esta discusión sobre la evolución convergente demuestra todavía más que la evolución es repetible en ciertas direcciones o vías importantes. Cuando la selección natural afecta de una misma manera a grupos lejanamente relacionados, frecuentemente encontramos resultados similares. En muchos casos, estos resultados similares son favorecidos por la historia anterior (homología) y se logran a través de rutas similares, aunque no idénticas, demostrando que la contingencia y el azar también están presentes. La evolución es, pues, un equilibrio entre sucesos contingentes, como las mutaciones y otros sucesos aleatorios, y otros claramente no contingentes, como la selección y la evolución convergente.
En el siguiente artículo de esta serie volveremos sobre la ecolocación de los murciélagos para explorar cómo la evolución de una especie puede ser en gran parte moldeada por otra especie en estrecha relación con ella (un fenómeno conocido como coevolución).
Notas
1.- Nota del T.: En el original ““The eyes have it”.
2. - Nota del T.: En el original, “Hearing is believing”
3. - Nota del T.: En el original “toothed whales” ballenas con dientes, es decir los odontocetos como los delfines, las orcas, etc.


